
苦草属(Vallisneria)植物的生态学研究概况
沉水植物作为主要初级生产者,在水生生态系统中有着不可替代的作用。当沉水植物丰富时,水体表现为水质清澈、溶解氧高、藻类密度低、生物多样性高等特点。因此,富营养化水体中沉水植物的恢复与重建已成为水域生态学研究的重点内容之一。苦草(Vallisneria)是淡水湖泊中常见的一种沉水植物。本文探讨了苦草属植物的生态学研究概况。
一、种属与分布
1.1 种 属
恩格勒系统(1964)
被子植物门 Angiospermae,单子叶植物纲 Monocotyledoneae,沼生目 Helobiae,水鳖亚目 Hydrocharitineae,水鳖科 Hydrocharitaceae,苦草属 Vallisneria linn:本属全球范围内有10种,我国有3种,南北各省区均产。
1.2 植物特征


1.3 苦草的生物分布
苦草属植物广泛分布于世界各地,尤以热带、亚热带和暖温带多见。在我国苦草属植物广泛分布于南北各省区,在淡水湖泊、沟渠、池塘、内河航道的静水或流动水体中均能很好地生长。密刺苦草是广东江段的建群种,长势好,蕴藏量大;而广西江段虽然还有密刺苦草,但其优势地位已逐步让位给亚洲苦草(V. asia -tica) ;到了云南江段却是苦草(V. natans)占据优势。目前在湖北洪湖和武汉东湖也先后报道了该种的存在。
刺苦草是长江中下游湖泊的优势物种,苦草为常见种,而密刺苦草(V. denseserrulata)仅有很少分布,苦草和刺苦草常混生于这些湖泊中,形成共存分布格局:刺苦草和苦草种间呈明显的的带状相间分布格局,或苦草只零星分布于刺苦草群落中。
二、繁殖特征及生活史
2.1 生长季节及温度
苦草是多年生草本。分布在亚热带地区的苦草( V. natans )冬季地上部分死亡,以冬芽或种子越冬,春季从冬芽或种子开始萌发,且随气温上升开始缓慢生长,夏季为快速生长期,最大生物量一般在9月份出现。开花期是5~ 8月,结果期为8~10月,种子成熟期在12月左右。

2.2 繁殖方式
苦草的繁殖方式:分为有性繁殖(种子繁殖)和营养繁殖(冬芽和活体植株)。
1、有性繁殖
植物产生有性生殖细胞(配子),经过两性生殖细胞的结合,成为合子,在由合子发育成为新个体(新的植株)的生殖方式,叫植物有性繁殖。
苦草具有强大的有性繁殖能力,苦草为雌雄异株,雄花在水面借助于空气和水流扩散时可能遇到雌花,当被水波暂时淹没时雄花可能被捕获到雌花花被里面而完成受粉过程。
1)繁殖选择
水深、风浪、光照、气候环境,营养状况对苦草对繁殖方式选择的主要影响因素。
2)授粉
苦草分为雌株和雄株,雄株授粉完成后就会死亡,雌株多年生。水媒传粉。
3)结果
在传粉以后把幼果拉入水中,在水面下约30cm处比较稳定的环境里生长(25~ 30℃)。
4)萌发
萌发率为 40% ~ 69%,天然土效果最好。 萌芽率受温度、种子保存时间和光照的影响,不受水质的影响。

2、营养繁殖
植物营养繁殖是指由植物体的根、茎、叶等营养器官或某种特殊组织产生新植株的生殖方式,属于广义的无性生殖范畴。
一般而言,无性繁殖方式在苦草生活史中起着更为重要的作用,冬芽和活体植株是苦草属植物最重要的无性繁殖器官,苦草在水底地面以无性繁殖方式迅速发展可以快速占据更广大的生境,以从基质中获得更多的营养。
苦草的营养繁殖受以下几种因素影响:
1、种群密度:在风浪较大的湖泊中恢复苦草种群应主要利用冬芽或鲜体,种子只适用于相对静止的浅水池塘种植。种植必须达到一定的种群密度。经调查太湖苦草冬芽平均90~ 226个/m2。
2、光照:块茎发芽对光照需求不大,但温度对块茎的发芽率有较大影响,20℃时发芽率最高,达90.2%± 12.6%,光照对幼苗生长有一定影响。苦草的块茎苗初期的伸长生长主要依赖于基部的根茎生长。
3、种植深度:苦草球茎的萌发受土层厚度的影响很大,被5cm土层覆盖时萌发率为100%;土层厚度为15cm时萌发率为73.3%;土层厚度为25cm时萌发率仅为3.3%。
4、机械损伤:苦草叶片生长最活跃的部位是在叶片下端2cm处,即绿色组织与白色组织的过度区,只要不破坏叶片基部的生长点,苦草的再生就非常迅速,当破坏了叶片的基部时,植株将逐渐死亡。


冬芽繁殖 活体植株
三、氮磷营养盐循环
苦草对水体营盐的去除率随生物量的增加不断增大,当生物量达到 2380.00g时,去除率放缓。苦草腐败释放的磷大部分被底泥吸附,少量进入水体。草腐败分解具有阶段性,前36d分解迅速,以后分解速率显著下降。
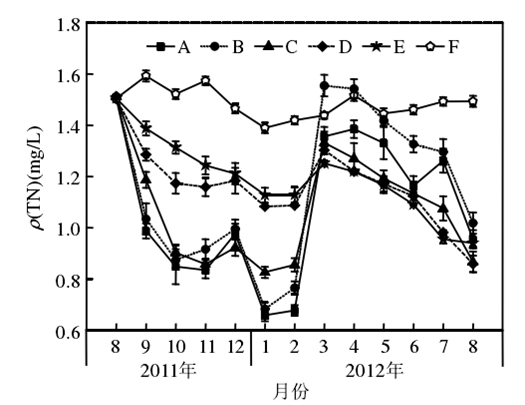
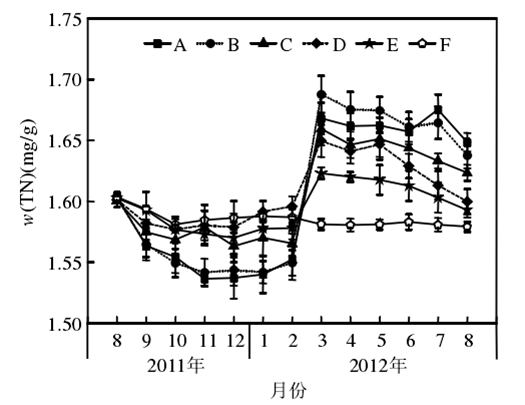
3.1 尿素氮的阈值范围
在富营养状态下(N:4 mg/ L;P:0.2 mg/ L)不同比例硝态氮(NO-3-N)和尿素氮(Urea-N)对苦草(Vallisneria atans)的生理影响:随着培养液中尿素氮比例的上升,生物量和硝酸还原酶活性,依次降低;POD和脲酶活性先升高后降低.研究表明,添加一定量的硝态氮可以促进苦草的生长,但当环境中尿素含量逐渐增加时会对苦草产生一定的胁迫作用,影响苦草的正常生长.从本实验研究结果综合来看,硝态氮(NO-3-N)和尿素氮(Urea-N)对苦草产生胁迫影响的阈值比为1∶ 1.此时TN为4 mg/ L,NO-3-N和Urea-N均为2 mg/ L。
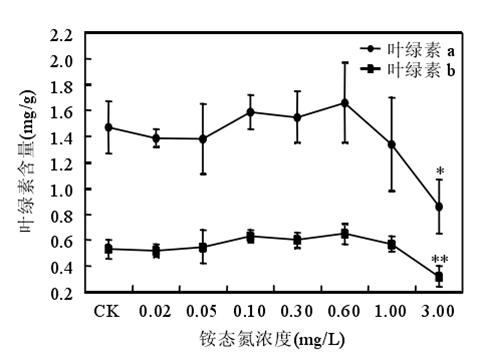
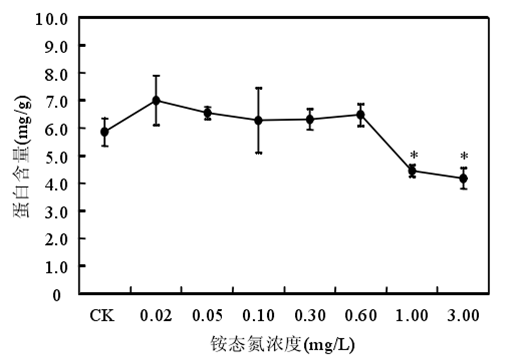
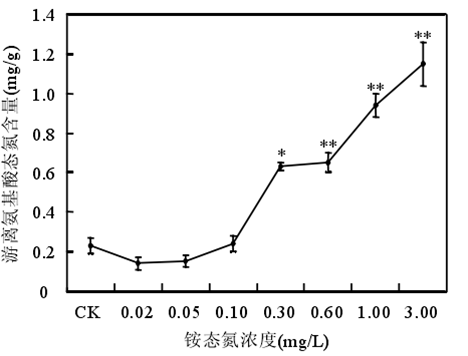
3.2 氨氮的阈值范围
铵态氮浓度低于 0.60mg/时苦草生长良好,浓度大于或等于 1.00mg/L 时苦草的光合能力受到抑制、代谢受到干扰。苦草对铵态氮最敏感的生理生化指标是叶片中游离氨基酸态氮含量。铵态氮作为沉水植物的一种营养物质,当其含量较低时,植物由于营养缺乏诱导产生自由基。
四、种群生态特征
4.1 生物量
在很多湖泊植被调查工作中都对苦草群落或含苦草的群落生物量进行了测定(见下图)。从表中可以看出,最高和最低生物量之间存在上千倍的差异。这一方面是因为调查对象尚包括其它种类,而非苦草本身; 另一方面,也是由于调查方法本身所致:
(1)调查结果往往用湿或鲜重表示。由于水生植物含水量丰富,离水后,因种类、天气、采样等因素的影响,其重量在短短的30 min内即能发生很大的变化。因此在对不同研究者的数据进行比较分析时,这便成为主要的误差来源。
(2)各研究中取样方法不规范,且各个方法的样方数和重复次数过少。样方重复次数过少常常导致结果存在极大的误差。因为从现有的研究看,群聚分布是我国水生植物最普遍的分布式样。
(3)测量时期不一致。一般各个研究均在最大生物量时期测量一次,但不同研究者对出现最大生物量时期看法并不一致,从6~ 9月均有,没有逐月的调查数据是难以得出准确的最大生物量出现时期的。
从部分湖泊的工作来看, 苦草生物量还随季节有明显的变化,同一湖泊不同地点以及同一湖泊不同年度苦草的生物量也有较大变化,但这些工作依然或多或少存在上述的取样问题。生物量的测定是研究植物生产力的重要基础,关于取样方法、样方数量、结果表达等方面国际上已存在被普遍接受的原则,为了准确评价水生植物的生产力,同时为了减少误差和进行国际交流,建议在今后的研究中应当以这些标准方法为指导。
(1).jpg)
4.2 群落特征
在我国多数水生植被调查工作中,群落的划分与命名一般采用优势度途径,而优势种的认定在许多情况下是以目测的表观优势度为标准,加之取样方法上存在的问题,多数工作缺乏严格的定量分析数据。
目前研究中涉及的不同群落仅仅是具体的地方群落( Local community) ,而并非群落分类学意义上的群落类型。根据近年来的研究,我们认为层片( synusia )途径是一个比较理想、实用的水生植物群落分类标准。利用层片分类系统,有可能对调查涉及的每一群落作出细致的分析。

五、个体生态特征
5.1 低溶解氧对苦草繁殖的影响
1、溶解氧平均值为1.6 mg/L时对苦草株重、株高、分蘖数及叶片数等指标的影响均不明显,对块茎的影响则较显著,表现为块茎数量与重量显著下降。
2、低氧对苦草根系的影响显著,表现为根须变细且数量增加,根系活力明显下降。
3、低氧处理使岸边沉积物处理组的沉积物氧化还原电位显著下降、水体营养盐浓度上升,尤其是磷酸盐浓度显著增加。低氧对苦草生长的影响虽不明显,但对其种群扩张有潜在的不利作用。
.jpg)
5.2 水体的营养水平对苦草生长的影响
1、随着营养盐含量的升高,苦草生物量的增长率逐渐降低,当水中ρ(TN) 达到10 mg/L和ρ(TP) 达到0.4 mg/L时,苦草的生物量开始减少;营养盐含量升高对苦草叶片特征的影响不明显,而苦草根状茎的生物量却随着营养盐含量的升高而逐渐减少。
2、当水体营养水平达到ρ(TN) 为10 mg/L和ρ(TP) 为0.4 mg/L时,除叶片长度外,苦草的其他形态指标值均显著下降。
3、当水中磷含量增加1 倍时对苦草生长造成的抑制作用大于氮含量增加1倍时;当二者均增加时,对植物生长造成的抑制作用显著增加。

5.3 水深光照的影响
1、苦草的水深分布
洱海苦草定植的水深范围为 0.5~5.6 m,抚仙湖苦草生长的最大水深为20m。鄱阳湖水淹深度7.2m 以内时冬芽萌发率和水深相关不密切, 超过 7.2m 以上时呈明显的负相关。
.jpg)
苦草分布湖泊的水质状况
2、光照需求范围
1)苦草幼苗正常生长发育所需的最小相对光照强度为1.6%。
2)弱光照和高营养均能对苦草生长产生抑制,两者有交互作用,主要是弱光影响高营养的抑制作用。水下光照低于10%水面光照后,弱光照成为苦草生长的主要限制因子。
3)水下光强不足自然光强的4.5%,幼苗叶片出现发黄、折断现象,幼苗光合作用受到明显抑制。水下的光强为水面光强的43.3%以上,苦草幼苗也受到抑制,光强范围大约是7.1%~43.3%。
(1).jpg)
.jpg)
5.4 水流对苦草的影响
1、苦草在静水水体中普遍存在,其强大的根系和柔软的带状叶使它能很好地生活在流动的水体中,即使在流速较大的溪流生境中也能生存。野外调查发现流速达到 0. 5 m•s- 1以上的生境中都能很好地生存。
2、为了更好的了解水流对苦草的影响,有研究人员层用水槽进行模拟流水,得出以下结论:
(1) 冠层以上,水流不受苦草冠层的影响,时均流速呈对数分布; 冠层内部,由于冠层阻流面积在垂线分布上存在差异,冠层内时均流速呈逆梯度分布,且在床面附近出现局部流速最大值。
(2) 冠顶处,水流强烈的紊动掺混与流速切变使得雷诺应力在冠顶处达到最大值,并向水面与床底方向逐渐减小; 冠层内部,逆流速梯度的出现使得雷诺应力出现负值,并在冠层内部出现局部最大值。
(3) 切应力产生的剪切紊动使得紊动能在冠顶附近最大,并向冠层内部进行垂向传输。受传输距离的限制,冠层底部水流以尾流紊动为主,紊动能大大减小。
.jpg)
5.5 植物化感与矮化处理
1、苦草可分泌化感物质酚酸,主要组分为:苯甲酸、对羟基苯甲酸、对羟基苯乙酸、邻苯二甲酸、对羟基苯丙酸、香草酸、原儿茶酸、阿魏酸和咖啡酸等九种酚酸。 酚酸抑藻测试的结果显示其抑藻活性与本身的结构有关,不同酚酸以毒性效应比例多维混合表现出加和抑藻效应,且随着混合种数的增多,酚酸的加和效应增强。
2、矮壮素2-氯乙基三甲基氯化铵浓度为 0. 02 g•L- 1左右时,对苦草矮化较为适宜。而矮壮素对自然条件下深水湖泊和开放性水体的生态影响,有待进一步研究。
六、思考与展望
1、目前关于苦草的研究主要集中在实验室阶段和野外调研,关于苦草的应用研究相对较少。
2、关于苦草的研究缺乏系统性和深度的研究,目前仍有较大的不确定性。
3、工程应用问题的解决必须建立在已有的基础之上,灵活运用自然规律和科学方法。盲目的进行开展工作,不仅会带来极大的资源浪费,还有可能徒劳无获。
参考文献:
[1]刘晓培,张饮江,李 岩,等.矮壮素对苦草矮化特征及生理指标的影响[J].生态学杂志 2012,31( 10) : 2561-2567.
[2]沈显生.安徽苦草属一新种[J].植物分类学报(2001)39(6):571~ 574.
[3]沈显生.安徽省苦草属种子的扫描电镜观察[J].暨南大学学报2001,22 (1 ):89-91.
[4]王春景 , 周守标 , 杨海军,等.安徽水生维管植物的多样性[J]南京林业大学学报2006,30(5)87-90.
[5]朱增银,尹大强,陈灿,等.不同比例硝态氮和尿素氮对苦草的生理影响 [J]南京大学学报(自然科学)2005,41(6):627-633.
[6]黄玉源,雷泽湘,何柳静,等. 不同磷浓度对水生植物苦草的影响研究[J]. 环境科学与技术, 2011, 34(2):12- 16.
[7]宋睿,姜锦林,耿金菊,等.不同浓度铵态氮对苦草的生理影响中国环境科学[J].2011,31(3):448~453
[8] 中国植物志编委会.中国植物志(第8 卷)[ M] .北京:科学出版社, 1992.176 ~ 181.


